摘要:种植单一的超富集植物修复重金属污染土壤,不但中断农业生产导致经济收益降低,而且因生物量较低、修复周期长等诸多弊端导致修复效果不甚理想。间作作为一种传统的农艺管理方式,利用生态位和生物多样性原理等能提高农作物对资源的有效利用,对共植的农作物种类增量提质。在中、轻度污染土壤修复中利用间作体系,通过调控超富集植物与农作物的生长发育,促进超富集植物根系低分子量有机酸(LMWOAs)的分泌,降低其根际土壤pH,增加重金属活性,从而增加超富集植物对重金属的吸收,同时抑制农作物根系LMWOAs的分泌,以减少农作物对重金属的吸收,提高其产量和品质,实现“边生产边修复”,提高土地利用率,并增加经济效益。本文根据近几年来国内外相关文献,综述了间作条件下超富集植物和农作物生物量、生理生化响应、重金属吸收、转运、富集等方面的变化,以及间作对土壤环境质量的影响,并对间作修复重金属污染土壤领域的发展趋势,如超富集植物和农作物间作的信号转导和分子生物学机制、间作体系下两类植物根际微生物类群的差异及其功能机制,以及构建高效间作体系提高重金属污染土壤的修复效率等方面进行了展望。
关键词:超富集植物;间作;重金属;农作物;土壤修复
当前我国所要解决的生态危机不是单方面的环境污染,而是人口激增、资源破坏和短缺等多方面综合效应。植物修复(phytoremediation)一般所采用的方法是利用超富集植物超强的重金属富集能力,吸收土壤重金属后及时收获植株,从而降低土壤中重金属含量[1]。然而超富集植物在修复过程中往往存在中断农业生产、生长周期长、生物量低等缺点,加之目前发现的重金属超富集植物往往具有经济价值低的特点,单一种植超富集植物难以激发农民的种植积极性,且当前农作物种植中,品种单一化不仅导致病虫害严重,而且杂草丛生,大量化学物质投入也造成土壤、水体和农产品的严重污染。在环境保护过程中,应以有限的资源为基础,生产出更多的产品,以满足经济和社会的发展需要,力求达到生态、社会和经济效益的协调统一。因此,为达到农业生产和土壤重金属污染治理的双重目的,做到“边生产边修复”,因地制宜建设良性的社会-经济-自然复合生态系统,超富集植物与农作物的间作修复应运而生。除了合理利用资源、获得良好的经济和社会效益外,还能促进农业生态环境的保护和修复。在超富集植物-农作物-土壤构成的间作体系中,各组分究竟发生了什么变化是一个值得深入总结和探讨的重要科学问题。本文综述了间作条件下超富集植物和农作物生物量、生理生化响应、重金属吸收、转运、富集等方面的变化,以及间作对土壤环境质量的影响,并对间作修复重金属污染土壤领域的发展趋势进行展望。
1重金属污染土壤间作修复的生态学意义
间作是我国精耕细作的农艺管理方式之一,利用人工调控物种共生原理,结构与功能相协调原则分层多级利用物质,对发挥土地生产力具有重要的生态经济价值。共生期内的两种作物种植可以增加同一地块的生物多样性并调节种间关系,通过协调种植过程中的物种配置和组合关系,使每种作物或品种在空间和营养生态位上互补扩大[2],实现作物对光、气、热、水、肥等资源的最大限度有效利用,减少病虫害的发生,同时增加土壤中一些酶的活性进而使土壤微生物生存环境得到改善,最终提高农作物产量和品质[3-4]。
在植物修复过程中应用间作体系,具有显著的生态学意义。通常采用高的农作物与矮的超富集植物间作,高农作物[如玉米(ZeamaysL.)等]为超富集植物提供了遮荫环境,在充分利用生态位的同时改善了超富集植物生长的小气候,增加物种多样性并减少病虫害的发生。此外,待修复完成后,及时收获富集了大量重金属的超富集植物并进行妥善处理,农作物的根和茎叶腐烂后,可以增加土壤有机质的含量,从而提高土壤肥力。总之,在种植超富集植物对土壤进行修复的过程中,通过与农作物间作进行可持续的农业生态系统设计,不需要停耕原来种植的农作物,可实现边生产边修复,从而提高了土地利用效率,产生显著的经济收益。玉米和铅超富集植物土荆芥(ChenopodiumambrosioidesL.)[5]、镉/锌超富集植物伴矿景天(SedumplumbizincicolaX.H.GuoetS.B.Zhousp.nov.)与玉米[6]、镉超富集植物龙葵(SolanumnigrumL.)与大葱(AlliumfistulosumL.)[7]等众多间作试验结果均表明,间作在不影响农作物长势、生物量的情况下,对土壤进行了修复。间作体系在强化植物修复效率的同时未损害农民的经济收益,并降低了土壤中重金属的含量,具有较强可行性与公众可接受性。
2间作修复条件下的超富集植物
2.1间作对超富集植物生物量的影响
超富集植物修复重金属污染土壤的原理是利用其根系选择性吸收重金属并运输至地上部,土壤所去除的重金属总量是植物组织的金属含量乘以生物量的积,较高的生物量意味着土壤中较多重金属被去除,取得更好的修复效果[8-9]。
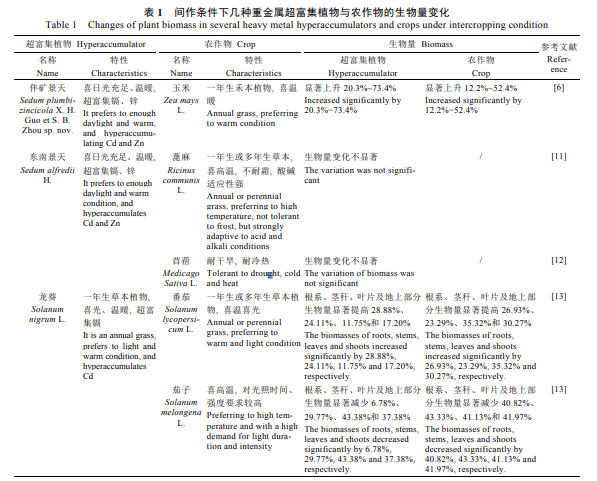
农作物类别和品种对超富集植物的生长发育具有促进与抑制的双重作用。在砷超富集植物大叶井口边草(Pteriscreticavar.nervosaThunb.)与不同品种玉米[10]、锌/镉超富集植物东南景天(SedumalfrediiH.)与蓖麻(RicinuscommunisL.)[11]、苜蓿(MedicagosativaL.)[12]、镉(Cd)富集植物龙葵[13]与茄子(SolanummelongenaL.)间作系统中,超富集植物的生长均受到抑制,其中与茄子间作的龙葵根、茎、叶及地上部生物量较龙葵单作显著减少6.78%、29.77%、43.38%和37.38%[13]。产生这种现象的原因可能是由于此类农作物与超富集植物对资源的需要或利用方式相同,形成了种间竞争,而农作物的竞争能力更强,可以更多地利用土壤中的氮、磷和有机质等,超富集植物在竞争中则处于劣势地位,导致其生物量显著降低[13]。但是,也有一些相反的结果。铅超富集植物土荆芥和蚕豆(ViciafabaL.)、玉米间作,伴矿景天与玉米间作[6],都不同程度提高了超富集植物的生物量,例如土荆芥地上部和根部生物量分别显著增加166%和134%。几种重金属超富集植物与部分农作物间作后的生物量变化情况如表1所示。从表中可以看出,选择生长习性合适的超富集植物与农作物间作、产生较高生物量从而更好地进行重金属提取对于间作体系至关重要。
将4个镉超富集植物孔雀草(TagetespatulaL.)的不同品种(矮秆红花、矮秆黄花、高秆红花和高秆黄花)与油菜(BrassicachinenesisL.)[19]间作后,发现超富集植物的根系和地上部分的生物量较单作显著增加,增幅分别为57.4%~89.9%、39.2%~57.5%。砷、汞胁迫对超富集植物生长都表现出一定的“低促高抑”作用,当砷和汞浓度累积到一定值时,会扰乱植物体内正常的新陈代谢,导致植物叶片坏死和枯萎、生物量降低甚至死亡[20]。间作所增加的植物地上部生物量能为重金属提供更多的蓄积位置,避免重金属过多聚集在某一部分,从而降低对植物生理生化造成的负面影响[21]。
2.2间作对超富集植物生理生化的影响
植物体内的活性氧(reactiveoxygenspecies,ROS)与抗氧化酶如超氧化物歧化酶(superoxidedismutase,SOD)、过氧化物酶(peroxidase,POD)和过氧化氢酶(catalase,CAT)等的活性之间存在动态平衡,是植物及时清除体内ROS并保持正常生长的内源保护机制[22]。然而重金属胁迫下,ROS在植物细胞内的过度积累将打破这种平衡从而导致蛋白质变性、DNA损伤、细胞酶失活以及膜脂过氧化损伤等,甚至会导致细胞死亡[23-24]。与番茄(SolanumlycopersicumL.)[13]间作后,镉超富集植物龙葵叶片SOD活性与单作相比无显著差异,但POD和CAT活性分别显著提高68.71%和5.66%。SOD催化超氧阴离子自由基歧化生成O2和H2O2;POD能调控植物体内吲哚乙酸(IAA)含量水平,免除机体内产生H2O2的毒害作用;CAT则将H2O2转化为H2O和O2,参与谷胱甘肽-抗坏血酸循环,从而将细胞中的H2O2去除[25]。间作体系中超富集植物体内抗氧化酶活性的增加降低了ROS的含量,缓解重金属带来的植物细胞过氧化程度,减轻了过氧化作用,从而在一定程度上缓解了细胞膜结构和功能的损伤,提高植物对重金属胁迫的耐性。
可溶性糖和可溶性蛋白在植物适应重金属胁迫的渗透调节中发挥着重要作用。龙葵与番茄间作[13]中龙葵叶片的可溶性蛋白含量显著增加,植株根系、茎秆、叶片及地上部分可溶性糖含量较龙葵单作分别显著提高89.06%、82.30%、79.76%和81.77%。这些结果表明,间作通过增强抗氧化酶活性、可溶性蛋白和可溶性糖含量,提高了超富集植物自身对重金属的耐受程度。
2.3间作对超富集植物富集重金属的影响
间作体系影响超富集植物对重金属的富集和转运。间作桑树(MorusalbaL.)[26]、构树[Broussonetiapapyrifera(L.)Vent.][17]提高了蜈蚣草(PterisvittataL.)体内砷的积累;在镉胁迫下,龙葵间作葡萄(VitisviniferaL.)幼苗[27]、牛膝菊(GalinsogaparviflorCav.)间作生菜(LactucasativaL.)[28]能显著增加超富集植物体内的镉含量。通过提高重金属在土壤中的生物有效性促进超富集植物地下部的吸收和地上部的积累,玉米间作使镉富集植物续断菊(SonchusasperL.Hill)[29]植株中镉含量提高,各镉浓度处理下(0~50mg·kg–1、200mg·kg–1)间作与单作续断菊相比,间作续断菊体内镉含量显著提高31.4%~79.7%,这与黑亮等[30]对间作玉米增加东南景天锌含量的结果相类似。这些结果表明间作体系提高了超富集植物对重金属的吸收和富集系数(植物体内重金属含量与土壤中重金属含量的比值),因此间作体系能更好地增加超富集植物对重金属的富集,提高土壤修复效果。
由于外部形态及内部结构的差异,植物吸收重金属元素的生理生化机制不同,不同部位对重金属的富集量也存在显著差异。从拔节期到成熟期,间作续断菊[31]的根部及地上部镉含量分别增加16.88mg·kg–1、15.45mg·kg–1,但单作仅分别增加5.5mg·kg–1和10.09mg·kg–1,两种不同种植方式的镉含量增幅存在显著差异。根是与土壤或水体直接接触的器官,作为吸收土壤中重金属的主要部位,根系状态(例如根长、根表面积、根体积、侧根数和根系活力等)影响着超富集植物吸收土壤中养分和重金属的能力。与单作相比,间作续断菊地上部和根部生物量、根长、根内径和根系体积均显著增加[32]。提高超富集植物根系发达程度有助于超富集植物吸收更多营养物质,增加更多的重金属储蓄位置从而增加重金属富集量。根系对土壤重金属的吸收能力较强但生物量较低,地上部拥有大的生物量意味着重金属含量的增加即更高的修复效率。
相关知识推荐:土壤有机质生态效应论文如何选刊发表
针对超富集植物间作后重金属含量与单作时存在差异这一结果,研究者用间作后的根际微环境变化来解释相关机理。秦丽等[32]发现间作增加了续断菊对铅的吸收和富集量,原因是间作体系中超富集植物根系分泌的低分子量有机酸(LMWOAs)增加了土壤有效态铅含量。续断菊根系所分泌的低分子量有机酸如柠檬酸是三元酸,具有良好的配位(络合)作用,能活化土壤中的重金属从而提高Pb2+活度。当两种植物间作时,超富集植物根系柠檬酸、草酸的分泌量增加,进而活化铅并促进对铅的吸收和富集。这与铅超富集植物小花南芥(ArabisalpinaL.var.parvifloraFranch)、锌/镉超富集植物东南景天地上部分铅、锌含量在间作条件下均增加的研究结果相似[33-34]。对于间作条件下农作物正常生长、超富集植物对重金属的提取能力增加,还可能与超富集植物对离子吸收运输的通道密切相关。例如Sasaki等[35]克隆出的耐铝基因ALMTI可以编码苹果酸转运子,在植物体内表达从而增加苹果酸的分泌量,活化重金属从而增加超富集植物的富集量。不同重金属胁迫、植物种类以及所分泌的各种低分子量有机酸对植物产生各种影响,表明有机酸分泌可能与离子通道有关。了解植物根系分泌通道与有机酸分泌的差异,对提高超富集植物对重金属的富集、降低农作物对重金属的吸收和转运,最终提高农作物的品质具有重要的现实意义。
3间作修复条件下的农作物
3.1间作对农作物产量的影响
产量是反映植物生长状况和经济收益的一个重要指标。对与镉超富集植物间作的十字花科(Brassicaceae)蔬菜的研究结果表明,与单作相比,水菠菜(IpomoeaaquaticaForsk)[36]根、茎、叶生物量分别显著增长28.72%、45.90%和7.74%,且根茎比(根生物量与茎生物量之比)也比单作提高。生菜与矿穗生态型牛膝菊(矿山生态型牛膝菊为接穗进行嫁接)间作[28],既能显著降低生菜可食用部位镉含量,还能促进其生长。在间作体系中蔬菜作物的地下部和地上部生物量均有增加,显著提高了作物的产量进而增加了经济价值。龙葵与玉米[37]、续断菊与玉米间作后,间作玉米籽粒产量显著高于单作,且与续断菊间作的玉米籽粒产量的收获指数与单作相比具有显著差异[31]。
植物通过光合作用合成有机物,是农作物产量形成的基础。间作之所以能够提高玉米的产量,是因为间作存在的植物株高差能改变冠层内的光分布并增加光能利用效率,从而增加叶片光合能力和干物质积累并分配到籽粒中[38-39]。间作体系中冠层内的小气候环境,特别是太阳辐射或光能利用效率对作物生产力至关重要,玉米需要更多光能,为其下生长的砷超富集植物蜈蚣草创造了喜阴环境,从而加快了蜈蚣草的生长发育。Singh等[40]发现砷、汞能取代叶绿素分子中的镁离子并降低叶绿素合成酶的活性,使叶绿素合成降低,同时增加叶绿素分解酶的活性,使叶绿素含量减少,从而降低了光合色素的含量和比例,光合参数的变化反映了光合作用的强弱,从而影响有机质的合成。Tang等[36]发现与单作相比,间作水菠菜的叶绿素a、总叶绿素和类胡萝卜素含量均显著增加,间作缓解了重金属带来的抑制,在加强农作物自身抵抗力的同时增加干物质的积累。
间作体系中作物生物量的变化是地上部与地下部综合作用的结果,且地下部作用大于地上部。间作小花南芥[33]使玉米分根条数、根表面积和根密度与单作相比分别显著增加60%、15%和42%,干重增加108%。作为固定、运输和吸收营养物质的重要器官,发达的根系能提高植株对土壤中营养物质的吸收利用,从而增加作物的生物量。
3.2间作对农产品品质的影响
不同间作方式下玉米籽粒砷积累水平均较低,能满足《食品安全国家标准食品中污染物限量》(GB2762—2017)中玉米籽粒砷含量限值[16]。谭建波等[31]认为间作增加了农作物的生物量,更大的生物量意味着重金属在籽粒浓度上产生了所谓的“稀释效应”,这与Feil等[41]、夏海勇等[42]发现氮肥施用提高玉米籽粒产量、降低重金属吸收量的结果相似。但在蜈蚣草与蓖麻[18]共植中,虽未对蓖麻产量产生显著影响,却降低了蓖麻籽粒的砷含量。这是由于间作使作物砷从营养器官向籽粒的转移速率受到抑制,从而籽粒的砷含量低于单作玉米。在单作或间作方式下,玉米吸收镉的主要部位均在根部,但间作显著降低了玉米的转运系数(植物地上部重金属含量与地下部重金属含量的比值),降低了玉米向地上部转运重金属的能力,所以籽粒中镉含量降低[29]。关于转运系数的变化可以从植物根系之间的“根际对话”研究得到解释。农作物体内重金属含量降低的一个主要原因是土壤胶体、土壤有机质特别是超富集植物的根系分泌物等吸附、固定了一部分游离的重金属。Yang等[43]发现一些植物分泌的磷酸盐可与铅结合生成难溶的Pb3(PO4)2,使土壤中的铅被钝化,降低了铅的生物有效性和移动性;间作系统中植物根系之间的相互作用进一步改变了根系分泌低分子量有机酸的组成和比例,超富集植物抑制了农作物根系分泌的低分子量有机酸的数量和类型,致使土壤中更少的重金属被活化,从而降低了农作物对重金属的吸收。间作续断菊体系发现玉米根系柠檬酸含量降低,更少的Pb2+被活化,根际土壤有效态铅含量降低,转运系数平均为0.56,显著降低12.51%[32]。另一个原因是超富集植物对重金属的吸收能力强于农作物,其根系能优先吸收和转运土壤中的重金属,减少了农作物根际土壤的重金属含量,从而降低主栽农作物对重金属的吸收。龙葵与玉米间作体系中[15],玉米根际镉含量和转运系数均显著降低,可食部分(籽粒)中的重金属含量显著减少,也说明由于根系间的一系列作用使植物根际环境发生变化,超富集植物发生类似向肥性的重金属“觅食”现象,抑制了玉米对重金属的吸收,通过对人体每天摄入镉量的安全值计算得出间作后土壤中镉全量低于2.26mg·kg−1,土壤有效态镉含量低于0.88mg·kg−1。该污染农田在间作模式下,食用玉米镉暴露对居民健康不存在风险。总体而言,间作系统能有效降低土壤中重金属含量,且收获作物的品质符合国家相关食品中污染物限量标准。——论文作者:郭思宇,王海娟,王宏镔
转载请注明来自:http://www.lunwencheng.com/lunwen/nye/19855.html
