摘要:本研究以1986—2016年珠江口海域游泳动物调查资料为基础,统计分析了近30年珠江口海域游泳动物经济物种群落结构变化特征及主要经济物种渔获率之间的相关性分析。结果表明:2000年以前和2000年以后珠江口海域游泳动物经济物种群落结构差异明显,2000年以前游泳动物经济物种种类数量显著高于2000年以后(F=6.711,n=1,p=0.017<0.05)。2000年以前主要游泳动物经济物种有蓝圆鲹(Decapterusmaruadsi)、蛇鲻(Sauridafilamentosa)、金线鱼(Nemipterusvirgatus)、鲐(Scomberjaponicus)、银鲳(Pampusargenteus)、灰鲳(Pampuscinereus)以及头足类,其中蓝圆鲹、蛇鲻、金线鱼等10种物种渔获率之间表现为显著相关(p<0.05);2000年以后的主要经济物种有周氏新对虾(Metapenaeusjoyneri)、脊尾白虾(Palaemoncarinicauda)、日本蟳(Charybdisjaponica)、直额蟳(Charybdistruncata)、凤鲚(Coiliamystus)、锐齿蟳(Charybdisacuta)、亨氏仿对虾(Parapenaeopsishungerfordi)、口虾蛄(Oratosquillaoratoria),主要渔获率均表现为显著相关(p<0.05)。2000年以前与2000年以后仅有4种优势经济物种相同,游泳动物经济物种群落结构发生了明显变化。
关键词:海洋生物学;游泳动物;经济物种;群落结构;渔获率;珠江口海域
河口是许多重要渔业资源的产卵场、索饵场和育幼场,为入海繁殖和溯河回游生物提供栖息地和洄游通道[1],同时也是重要的经济捕捞区,为全球提供大约25%的初级生产力,并支持全球90%的捕捞业[2]。渔业资源群落结构作为一个重要空间尺度生态单元,反映了渔业生态系统的物质循环和能量流动[3]。目前,国内外已有大量关于渔业资源随时间变化的研究。在国外,Bender等(2013)报道称以前渔民捕捞的鱼比现在体型更大,表明巴西东北部累西腓海岸公园中大型鱼类数量正在减少[4];Vinni等(2019)研究结果表明,随着时间的推移,河口渔获量高的渔区数量正在减少[5]。而在国内,Jin等(2013)、Chen等(2015)、Shan等(2016)研究表明由于受到过度捕捞、气候改变和水质污染的影响,黄河口渔业资源群落结构变化明显和物种多样性显著下降[1-3]。
海洋渔业资源长期以来一直是人类蛋白质摄食的主要来源之一,为人类社会发展提供了源源不断的优质水产品[6-7]。珠江口海域处于亚热带海域向热带海域的过渡地带,珠江径流挟带的大量营养盐入海使珠江口海域成为我国近岸海域最具生产力的水域之一。珠江口渔场是我国南方著名的海洋渔场,海洋渔业资源丰富,游泳动物群落结构特征十分复杂[8-9],其中游泳动物经济物种是构成海洋渔业资源的关键生物类群[10]。近30年来,随着珠江三角洲区域社会经济快速发展,各种人类活动对海洋渔业资源影响强度日趋加大,游泳动物经济物种群落结构在逐年变化。国内相关学者一直在持续关注珠江口渔业资源群落结构变化,并开展了间断性的珠江口海域游泳动物物种组成、群落结构调查研究[6]。这些研究多为单次游泳动物调查分析评估报告,缺乏对珠江口海域近30年游泳动物经济物种群落结构的整体变化特性剖析[11]。在当前我国尤其重视海洋环境保护和渔业资源养护的大背景下,全面分析珠江口海域游泳动物经济物种群落结构变化特征,可为珠江口海域游泳动物资源量恢复、海洋生态保护及海洋牧场建设等提供可靠的技术支撑[12]。
1材料与方法
1.1研究区域
珠江口海域处于粤港澳大湾区南部的南海近岸海域,东西宽度约为150km,面积约为6000km2,地理坐标为21°30'~22°32'N,112°50'~114°20'E,为粤港澳大湾区的海上屏障(图1)。珠江口海域盛产蓝圆鲹(Decapterusmaruadsi)、带鱼(Trichiuruslepturus)、大黄鱼(Larimichthyscrocea)、真鲷(Pagrosomusmajor)、青鳞(Sardinellazunasi)等,贝类资源也很丰富,藻类有马尾藻属(Scagassum)、紫菜属(Porphyra)和鹅肠菜属(Moench)等,水产养殖业发达。2015年珠海市万山海洋开发试验区的广东省万山海域获批第一批国家级海洋牧场示范区,示范区面积为312km2;2018年珠海市外伶仃洋海域的广东省珠海市外伶仃洋海域获批第四批国家级海洋牧场示范区,面积为9.83km2。
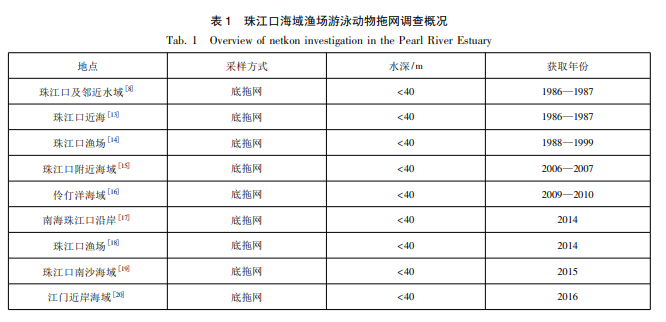
1.2数据来源与分析
本研究以近30年珠江口海域游泳动物调查研究资料为基础(表1),所选数据均根据《海洋调查规范》[21]和《海洋监测规范》[22]执行获得,且水深均小于40m。
2结果与分析
2.1珠江口海域游泳动物物种组成变化
对珠江口海域游泳动物调查的渔获物种类数量统计结果见图2。从图2可以看出,随着时间的推进渔获物种类数量明显下降(F=6.711,n=1,p=0.017<0.05),2000年以前渔获物种类数量明显高于2000年以后,其中1986—1987年渔获物种类数量最多,共有207种,2006年渔获物种类数量最少,只有28种。渔获物物种组成聚类分析表明,近30年珠江口海域渔获物物种组成显著分为两个时期,分别为2000年以前与2000年以后(图3),以2000年为时间界点,渔获物物种组成结构发生了显著变化。基于物种优势度、生物量占比和渔获率对2000年以前和2000年以后的物种组成进行对比分析,如表2所示,结果表明2000年以前优势种有丽叶鲹、黄斑鲾、杜氏棱鳀等13种,2000年以后优势种有深水金线鱼、棕斑腹刺鲀、中华青鳞等12种;2000年以前和2000年以后游泳动物优势种仅有4种相同,分别是黄斑鲾、棘头梅童鱼、中华青鳞和棕斑腹刺鲀;2000年以前生物量占比超过10%的有丽叶鲹、棘头梅童鱼和凤鲚,2000年以后生物量占比超过10%的仅有鲈;2000年以前渔获率超过10%的有丽叶鲹、蓝圆鲹、棘头梅童鱼和凤鲚,2000年以后渔获率超过10%的仅有康氏小公鱼。
2.22000年以前珠江口海域游泳动物经济物种的群落组成
基于主要渔获率群落组成进行物种之间的相关性分析,结果如图4所示。长尾大眼鲷与带鱼呈显著负相关(p<0.01,r=-0.73);头足类分别与金线鱼、鲐呈显著正相关(p<0.01,r=0.70,表示两两物种之间关系,并且数值相同,下同),金线鱼与鲐呈显著正相关(p<0.01,r=0.76);蓝圆鲹分别与丽叶鲹、银鲳、灰鲳、杜氏棱鳀、赤鼻棱鳀呈显著负相关(p<0.01,r=-0.75),蛇鲻分别与丽叶鲹、银鲳、灰鲳、杜氏棱鳀、赤鼻棱鳀呈显著负相关(p<0.01,r=-0.81);银鲳、灰鲳、杜氏鲮鳀任二者间均呈显著正相关(p<0.01,r=1.00),丽叶鲹分别与银鲳、灰鲳、杜氏棱鳀、赤鼻棱鳀呈显著正相关(p<0.01,r=1.00)。
相关知识推荐:期刊发表论文时间大约多久
对相关性高的物种进行生物学特性、感官评价、经济价值评估,进一步确定2000年以前游泳动物主要经济物种为蓝圆鲹、蛇鲻、金线鱼、鲐、乌贼、银鲳、灰鲳(表3)。
2.32000年以后珠江口海域游泳动物经济物种的群落组成
基于2000年以后经标准化的主要渔获率群落组成(图5),不同年份间主要渔获率差异很大:2006年渔获率占绝对优势的物种有周氏新对虾;2009年有脊尾白虾、日本蟳、棘头梅童鱼;2014年有棘头梅童鱼、直额蟳、黑尾小沙丁鱼、凤鲚、红狼牙鰕虎鱼;2015年有脊尾白虾、锐齿蟳、孔鰕虎鱼、短吻鲾、亨氏仿对虾;2016年有口虾蛄、隆线强蟹、太阳强蟹。对其主要渔获率群落组成进行物种间相关性分析,结果表明,2000年后渔获率占绝对优势的物种均表现为相关性高且极显著(p<0.01)(图6)。对以上渔获率占绝对优势的物种进行生物学特性、感官评价、经济价值的评估(表4),周氏新对虾、脊尾白虾、日本蟳、直额蟳、凤鲚、锐齿蟳、亨氏仿对虾、口虾蛄为2000年以后的主要游泳动物经济物种。
2.4讨论
2.4.1近30年游泳动物群落结构变化海洋游泳动物群落结构组成变化受海洋游泳动物存量、捕捞作业以及海洋环境变化等多种因素的影响。近30年珠江口海域游泳动物群落结构可以划分为2000年以前和2000年以后两个阶段。2000年以后的游泳动物经济物种种类数量较2000年以前有明显的减少。2000年以前游泳动物主要经济物种中,鱼类有6种,占85.7%;2000年后游泳动物主要经济物种中,鱼类仅有1种,占12.5%,主要经济物种从原来的经济价值高的大型鱼类开始向经济价值较低的鱼类和虾蟹类转化。这与谢志超等(2018)对珠江口游泳动物优势种组成结构的调查结果一致[20],一些经济价值高的鱼类物种优势度逐渐降低,被甲壳类所取代。山东南部近海主要游泳动物评估结果也与此趋势一致[27]。2000年之后与2000年之前相比,捕获的种类有明显差异,优势种游泳动物群落结构发生明显变化,造成这种变化的原因可能是:珠江口海区鱼类按生态类型分类时,中上层鱼类与整体资源种类数量变化趋势相同,2000年以前以中上层鱼类丽叶鲹作为绝对优势种,中上层鱼类优势地位的丧失,造成游泳动物种类的明显变化,这与莱州湾渔业资源群落结构表现的底层鱼类与整体资源种类变化趋势相同的结果有差异[28],推测南、北方游泳动物存在一定的差异,在制定管理策略时应因地制宜。
2.4.22000年以前与2000年之后主要经济功能群变化与2000年以前相比,随着珠江三角洲经济的高速发展,珠江口海域海洋生态环境受到陆源排污、海水养殖、气候变化等多种因素影响,赤潮等海洋生态灾害爆发频次明显增加,缺氧海域面积扩大[29],游泳动物生物群落生存环境劣化,导致生物种类明显减少。与2000年之前相比,游泳动物优势种发生明显变化,造成这种变化的原因可能是:高强度捕捞对游泳动物产生深远的影响,游泳动物经济物种优势种在高强度捕捞压力下难以恢复到正常水平[30],导致珠江口海域游泳动物生物量占比和渔获率均较2000年以前有明显的下降。一些经济价值高的鱼类如丽叶鲹、棘头梅童鱼和风鲚捕获比例锐减,渔获物组成倾向于低值化、小型化的鱼类。虾蟹类因生命周期短,适应能力和繁殖能力强,近年来的增殖放流和伏季休渔制度的实施,虾蟹类有取代上层优质鱼类成为主要经济种类的趋势。游泳动物经济物种结构的低值化在加剧,群落结构趋向于r-选择(有利于增大内禀增长率的选择称为r-选择,在一定意义上它们是“机会主义者”),1987—1998年捕获的游泳动物种类数量有一定程度的增加,这可能是由于捕捞强度和先进捕捞方式的应用造成的。2006—2016年间游泳动物经济物种种类数量虽然整体比2000年以前有明显减少,但在此期间游泳动物经济物种种类数量有缓慢增加的趋势,至2016年数量最多,这可能与1995年伏季休渔以来游泳动物恢复有关。
2.4.3游泳动物保护与管理虽然近年来,渔政管理部门已采取了许多游泳动物保护措施,如延长伏季休渔期、实施重要游泳动物经济物种增殖放流、划定水产种质资源保护区、开展现代化海洋牧场建设等。首先,伏季休渔虽然在保护游泳动物方面发挥了一定积极作用,但仍没有从根本上解决游泳动物衰退的问题,相关研究表明伏季休渔前后的捕捞强度差异明显,鱼类群落结构较单一[31]。伏季休渔期过后,游泳动物种类数量的下降和游泳动物主要经济物种群落结构的改变,以及鱼类在游泳动物经济物种群落结构中地位的下降,说明珠江口海域仍然存在较大的捕捞压力,即使休渔期适当延长,之后的连续高强度捕捞仍然对游泳动物恢复产生较大的压力[7]。其次,渔业管理需要以最大限度实现生态系统的综合效益为目的,相关研究已采用生态系统建模的方法探讨了最优捕捞政策,基于多物种模型的管理政策能清楚地解释渔业管理中经济、社会和生态系统目标之间的权衡[32]。生态、经济和社会效益之间的权衡是渔业管理的重要问题,尽管生态和社会经济目标之间存在着相反的关系,但这也使得在最大化系统整体的效益中找到平衡成为可能[33],为以后的研究提供方向。
3结论
本研究在系统分析近30年珠江口海域游泳动物调查数据的基础上,将游泳动物分为2000年以前和2000年以后两个时间段,2000年以前的游泳动物种类明显多于2000年以后的资源种类。2000年以前,游泳动物优势种有丽叶鲹、黄斑鲾、杜氏棱鳀等13种,其中蓝圆鲹、蛇鲻、金线鱼、鲐、头足类、银鲳、灰鲳为主要经济物种;2000年以后,游泳动物优势种有深水金线鱼、棕斑腹刺鲀、中华青鳞等12种,其中周氏新对虾、脊尾白虾、日本蟳、直额蟳、凤鲚、锐齿蟳、亨氏仿对虾、口虾蛄为主要经济物种,2000年以前和2000年以后的游泳动物优势种中仅有4种相同。2000年以前游泳动物经济物种中,鱼类有6种,占85.7%,2000年以后游泳动物经济物种中,鱼类仅1种,占12.5%,表明近30年珠江口海域游泳动物经济物种群落结构发生了明显变化。
因此,建议近岸海域捕捞应着重控制捕捞强度、调整捕捞方法;同时通过现代化海洋牧场建设等途径,加强近岸海域渔业资源养护和渔业生态环境保护,推动近岸海洋生态系统向多功能、健康、稳态、持续产出的方向发展。——论文作者:徐鹏1,3,谢木娇1,3,周卫国1,孙英婷1,丁德文1,2,索安宁1,2*
转载请注明来自:http://www.lunwencheng.com/lunwen/jgu/20085.html
