摘要H9N2亚型禽流感病毒(avianinfluenzavirus,AIV)在我国广泛流行,给养禽业造成了巨大的经济损失,还对公共卫生安全存在潜在威胁.2014~2016年,H9N2亚型禽流感病毒在江苏地区的活禽市场的鸡中的阳性率为8.39%~13.74%.选取10株H9N2分离株进行全基因组测序及遗传进化分析.结果表明,H9N2分离株的HA基因与早期的江苏分离株位于同一小分支上,且同源性较高,但NA基因与早期江苏病毒株的同源性较低,核苷酸同源性为91.4%~93.9%,氨基酸同源性为91.2%~93.8%.这些病毒株的基因型依然为B69,为目前的优势基因型.所有H9N2病毒分离株的HA蛋白的226位点为L.固相受体结合实验结果显示,H9N2分离株具有人源α2,6唾液酸受体的结合特性.
关键词禽流感病毒,H9N2,进化分析,基因型
禽流感(avianinfluenza,AI)是由正黏病毒科A型流感病毒(avianinfluenzavirus,AIV)引起的禽类高度接触性传染病,有16个HA亚型和9个NA亚型[1].根据禽流感病毒致病性的不同,可分为高致病性禽流感病毒和低致病性禽流感病毒.目前,我国的低致病性禽流感疫情以H9N2亚型禽流感为主.自1994年我国首次报道鸡(Gallusgallusdomesticus)群中分离到H9N2亚型AIV以来[2],该亚型禽流感病毒在我国范围内广泛存在,可导致产蛋量下降,与其他病原共感染时常导致高死亡率,给养禽业造成了巨大的经济损失[3~5].H9N2AIV也可直接感染哺乳动物,在1999,2003,2013和2018年,分别发生H9N2AIV感染人的事件(http://www.who.int/influenza/human_animal_interface/Influenza_Summary_IRA_HA_interface_24January14.pdf)[6~8].猪(Sus)是AIV“禽-猪-人”传播链中重要的中间宿主,是人流感病毒与AIV发生重组的搅拌器和放大器,H9N2AIV也可在猪中感染与传播[9].此外,H9N2AIV内部基因具有非常好的兼容性,可与其他亚型流感病毒发生重排,从而产生新型重排流感病毒,2013年H7N9亚型流感病毒和H10N8亚型流感病毒的内部基因就是由H9N2AIV提供的,其中H7N9病毒可导致大于25%的感染者死亡,在社会上引发了巨大的恐慌[10,11].
H9N2AIV为单股负链的RNA病毒,基因组分为8个节段,极易发生突变和重排.研究表明,H9N2AIV的基因型在进化过程中可呈现多样性,根据HA基因可分为7个分型,分别为A~G[12].2009~2012年期间,H9N2AIV可分为8个基因型(B64~B71),其中B69为优势基因型[13].定期分析H9N2AIV遗传进化及其分子特性,及时了解病毒变异和重组情况,对H9N2AIV的防控是非常必要的.受体结合是流感病毒感染和适应宿主的第一步,受体的特异性是流感病毒跨宿主传播的关键[14].之前已有学者发现,H9N2AIV通过HA蛋白发生Q226L的改变,其受体结合特性从主要结合禽源受体α2,3唾液酸受体转换为结合人源受体α2,6唾液酸受体[4,15].本研究在我国江苏省分离获得了10株H9N2AIV(2014~2016年),对其进行了全基因组测序,分析了其遗传进化,绘制了基因型,并分析了其分子特性,包括受体结合特性,为H9N2AIV的有效防控提供了依据.
1材料与方法
1.1病毒的分离
将从江苏省活禽市场收集的咽喉拭子样品(2014~2016年),加入1.0mL灭菌磷酸盐缓冲液(phosphatebuffersaline,PBS),5000r/min离心10min,取上清液,过滤除菌.将上述处理的上清液经尿囊腔接种9~11日龄SPF鸡胚,0.1mL/胚,弃去接种24h死亡的鸡胚,37℃孵育72h后置4℃过夜,收集鸡胚尿囊液.经血凝实验和国家标准中特异性H9特异性引物PCR检测为阳性的尿囊液,?80℃保存.江苏H9N2病毒株Ck/Jiangsu/825/2009,Ck/Jiangsu/925/2009,Ck/Jiangsu/C1020/2012,Ck/Jiangsu/C1243/2012和Ck/Jiangsu/A2093/2011为实验室保存[13].
1.2病毒的增殖与纯化
将上述病毒分离株,稀释后接种9~11日龄SPF鸡胚,通过有限稀释法纯化3次.纯化的病毒扩增后,分装作为种毒,?80℃保存.
1.3全基因组测序和遗传进化树分析
利用Trizol(购自TaKaRa公司,日本)提取H9N2亚型禽流感病毒各分离株的总RNA,反转录成cDNA,用文献中引物[16]进行8个片段的PCR扩增.PCR产物测序的结果用Seqman软件拼接,并用MEGA5.2软件的Clustal对比方法及neighboru-joining方法绘制了发育进化树,bootstrap值设置为500.各个进化树皆以Ty/Wisconsin/1966(H9N2)为根.分析的基因片段对应的位置分别为HA基因(256~1073bp)、NA基因(1~1377bp)、PB2基因(31~2258bp)、PB1基因(1~2208bp)、PA基因(1~2128bp)、NP基因(31~1438bp)、M基因(1~946bp)、NS基因(13~825bp).此外,用Lasergene软件(Dnastar,Madison,WI)中的MegAlign软件分析不同病毒株的核苷酸或氨基酸的同源性.
1.4固相受体结合实验
将Strepavidin-96-wellstripplates(购自Thermo公司,美国)用洗脱液洗3遍,3min/次;加入100μL的糖基3′SLN-PAA或6′SLN-PAA(由糖基学会提供),4℃过夜,然后用洗脱液洗3次,加入32U血凝效价的H9N2病毒液,4℃过夜;洗脱液洗3次,加入100μL鸡源H9多抗(自制,1:1000),4℃保持2h;洗脱液洗3次,加入100μL1:4000稀释的HRP-羊抗鸡(购自Sigma公司,德国)二抗,4℃保持2h;洗脱液洗3次,加入100μL邻苯二胺(ophenylenediamine,OPD)显色4min,加入50μL2mol/L硫酸终止液终止显色;OD492下读值.
2结果
2.1H9N2AIV分离
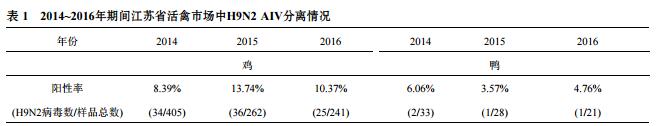
2014~2016年期间,在江苏省多个活禽市场的鸡或鸭(Anatinae)中共采集拭子1090份,其中鸡的样品为908份,鸭的样品为82份.H9N2亚型禽流感病毒主要分离于鸡,不同年代的H9N2阳性率为8.39%~13.74%,而在鸭的分离率会较低,不同年代H9N2阳性率为3.57%~6.06%(表1).
2.2HA基因遗传进化分析
为了进一步分析H9N2AIV遗传进化关系,挑选了10株2014~2016年期间分离于活禽市场的H9N2AIV分离株.在鸡胚上进行有限稀释纯化病毒,并进行全基因组测序.依据国内流行株及国内外参考毒株核苷酸序列的同源性,利用MEGA5.2软件绘制了HA基因的系统发育树(图1).结果表明,该10株H9N2AIV的HA基因皆属于Ck/BJ/1/1994分支下的SubgroupⅡ亚分支,而商品疫苗株Ck/Shanghai/F/1998位于Ck/BJ/1/1994分支下的SubgroupⅠ亚分支.本研究中,9株分离株的HA基因同源性很高,彼此的核苷酸的同源性为95.6%~98.8%,氨基酸同源性为95.7%~99.1%,聚在相近的分支上,且分布在早期江苏分离株Ck/Jiangsu/C1020/2012和Ck/Jiangsu/C1243/2012邻近,但有一株分离株Ck/Jiangsu/F345/2015分布于另一株早期江苏分离株Ck/Jiangsu/A2093/2011的附近,二者核苷酸和氨基酸的同源性均为100%.
2.3NA基因遗传进化分析
依据国内流行株及国内外参考毒株核苷酸序列的同源性,利用MEGA5.0软件绘制了NA基因的系统发育树(图2).结果表明,与大部分2009~2012年H9N2AIV一致,这些分离株的NA基因皆属于Ck/Shanghai/F/1998分支.进一步分析发现,这10株分离株的NA皆位于同一个小分支上,同源性很高,核苷酸同源性为96.4%~98.2%,氨基酸同源性为96.1%~97.9%.但与早期江苏分离株Ck/Jiangsu/A2093/2011,Ck/Jiangsu/C1020/2012,Ck/Jiangsu/C1243/2012和Ck/Jiangsu/DD6/2013同源性相对较低,核苷酸同源性为91.4%~94.0%,氨基酸同源性为91.2~94.2%.
2.4内部基因遗传进化分析
H9N2AIV具有非常好的兼容性,可为其他亚型流感病毒提供内部基因,从而形成新的重排病毒.10株H9N2AIV分离株的PB2基因属于Ck/Zhejiang/B2013/2012分支(图3A),核苷酸的同源性为94.8%~99.6%,氨基酸同源性为98.2%~100%,其中Ck/Jiangsu/F344/2015和Ck/Jiangsu/F345/2015PB2的氨基酸同源性为100%.分离株的PB1基因属于Ck/Shanghai/F/1998分支,9株病毒株的PB1聚在进化树的一个小分支上(图3B),1株病毒株Ck/Jiangsu/F345/2015与其他9株病毒株的遗传距离较远.PB1同源性分析比较结果显示,Ck/Jiangsu/F345/2015与其他9株病毒株的同源性略低,核苷酸同源性为95.1%~96.4%,氨基酸同源性为97.5%~98.8%.同期,这9株病毒株的核苷酸同源性为96.5%~98.8%,氨基酸同源性为98.2%~99.2%.这些分离株与2009~2012年的江苏分离株Ck/Jiangsu/825/2009,Ck/Jiangsu/925/2009,Ck/Jiangsu/A2093/2011,Ck/Jiangsu/C1020/2012,Ck/Jiangsu/C1243/2012和Ck/Jiangsu/DD6/2013之间的核苷酸同源性为95.6%~98.3%,氨基酸同源性为97.5%~99.5%.分离株的PA基因属于Ck/Shanghai/F/1998分支,其中3株Ck/Jiangsu/E1065/2014,Ck/Jiangsu/E135/2014和Ck/Jiangsu/E1158/2014聚在一个小分支上,另外7株病毒聚在另一小分支上(图3C),两者之间的核苷酸同源性为96.1%~96.7%,氨基酸同源性为98.9%~99.6%.NP基因属于Ck/Shanghai/F/1998分支(图3D),但其中3株Ck/Jiangsu/F344/2015,Ck/Jiangsu/F345/2015和Ck/Jiangsu/G236/2016之间的同源性较高,而其余7株间的同源性较高.分离株的M基因属于Ck/Zhejiang/B2013/2012分支,且聚集在一个小分支上(图3E).分离株的NS基因也属于Ck/Zhejiang/B2013/2012分支,但其中Ck/Jiangsu/E158/2014远离其余的9株分离株(图3F).
2.5基因型分析
在H9N2AIV的进化过程中,形成多种基因型.为了进一步了解该亚型病毒的基因型,根据分离株8个片段的系统发育树,绘制了病毒分离株的基因型(图4).结果表明,10株2014~2016年的江苏分离H9N2AIV的基因型皆为B69,与2009~2012年H9N2AIV的基因型相同.该基因型依然为目前国内H9N2AIV的优势基因型.
2.6分子特性分析
在本研究中,10株江苏省2014~2016年分离株在HA蛋白226位点上皆为L(表2).随机挑选其中的3株病毒株(Ck/Jiangsu/E135/2014,Ck/Jiangsu/E158/2014和Ck/Jiangsu/E1196/2014),用固相受体结合实验检测其受体结合特性.结果表明,该3株病毒皆表现为主要结合人源受体α2,6唾液酸受体的特性(图5).比较与受体结合相关氨基酸位点183和228位点,这10株病毒与较早分离的H9N2AIV一致,分别为N和G.与Ck/Jiangsu/825/2009毒株不同(含PSKSSR),分离株的HA裂解位点全部为PSRSSR.与疫苗株Ck/Shanghai/F/1998及2009~2012年的H9N2分离株一致,本研究中的H9N2AIV分离株的NA蛋白均缺失62~64位氨基酸(表2).研究发现,当M2蛋白为27V或31N时,流感病毒表现为金刚烷耐药性[17],而这些分离株的M2蛋白全部为27V和31N,表明这些毒株均为金刚烷耐药毒株.此外,这些毒株的PB2蛋白627和701位点均为禽源的E和D.
相关知识推荐阅读:动物检疫论文发表难度大吗
在论文发表过程中经常有作者问,论文发表难度大吗?其实论文发表难度大不大,并没有确切的答案,而是由多个因素决定的,动物检疫论文也是如此。有发表论文需求的作者,可以咨询我们在线编辑,给大家提供论文选刊、润色、修改、翻译、发表等服务。
3讨论
低致病性H9N2亚型禽流感病毒主要流行于欧亚等国家,与其他病原共感染时常导致高死亡率,给养禽业造成了巨大的经济损失[4].上海市动物疫病预防控制中心的一份2009年监测报告显示,在上海三大活禽批发市场中的样品H9亚型AIV平均阳性率为8.14%,在免疫H9N2亚型AIV灭活苗的抗体合格率达到100%的样品中分离到了8株H9亚型AIV(8/90)[18].2009~2012年H9N2AIV在全国多个省市活禽市场的分离率为8.20%~11.58%[13].相似的,在本研究中,活禽市场里的鸡携带H9N2AIV的阳性率达到8.39%~13.74%,但鸭子的H9N2AIV分离率低于鸡.
流感病毒的表面蛋白有两种,分别为HA蛋白和NA蛋白,在流感病毒感染与传播中起着重要作用,也是病毒的主要抗原[19,20].遗传进化分析结果表明,10株江苏省H9N2病毒(2014~2016年)的HA基因都处在早期的江苏H9N2分离株的小分支上,且它们的核苷酸和氨基酸的同源性都较高,说明分离株HA基因进化速度较慢.尽管如此,这些分离株和2009年后的大部分H9N2病毒的HA基因与疫苗株Ck/Shanghai/F/1998的遗传距离较远,在不同的亚分支上.分离株的NA基因与江苏早期分离株(Ck/Jiangsu/A2093/2011,Ck/Jiang-su/C1020/2012和Ck/Jiangsu/C1243/2012)同源性较低,表明NA基因在序列上变化较快,值得后期重点监测.但造成这一现象的原因尚不清楚,可能是因为该基因进化速率快,或者是因为其他亚型流感病毒的NA基因重排,需进一步深入研究.
自2009年开始,H9N2AIV流行株的基因型发生了较大的改变,主要为B69基因型[13].相较于以前的毒株,该基因型的H9N2病毒能更稳定地存在于自然界,且对鸡的致病性和接触传播性更强[5].研究表明,M基因是增强该基因型的H9N2AIV致病性的关键基因之一[21].本研究中的10株H9N2分离株的基因型并没有发生变化,仍然是B69.
禽流感病毒偏嗜α2,3唾液酸受体,人流感病毒偏嗜α2,6唾液酸受体.禽流感病毒的受体结合特性由α2,3唾液酸受体向α2,6唾液酸受体转变,是禽流感病毒感染人的关键.早期,H9N2AIV只感染水禽和鸡,其受体特异性为禽源的α2,3唾液酸受体.继1999年人感染H9N2AIV后,倾向结合人源α2,6唾液酸受体的H9N2AIV陆续出现[6].本研究中的H9N2AIVHA蛋白皆为226L,呈现为主要结合人源α2,6唾液酸受体的特性,固相受体结合实验也进一步证实该结论.
总之,H9N2AIV分离株的基因型依然未发生变化,B69为优势基因型,但近年来其受体结合特性转变为人源α2,6唾液酸受体,且NA基因序列变化也较大,可能对人和公共卫生的潜力危险进一步加大,需要加强对该病毒的监测.
转载请注明来自:http://www.lunwencheng.com/lunwen/yix/15680.html
